
|
Содержание сайта =>> Российское гуманистическое общество =>> «Здравый смысл» =>> 2007, № 2 (43) |
ЗДРАВЫЙ СМЫСЛ Весна 2007 № 2 (43)
ГУМАНИЗМ И ЭВОЛЮЦИОНИЗМ
Проблемы эволюции
|
Эволюция – evolution – в латинском языке обозначает «развёртывание». В биологии термин эволюция впервые применил Ш. Бонне (1762). Следует сказать, что уже в трудах античных философов Демокрита, Эмпедокла, Лукреция Кара и др. отражены представления о развитии жизни на Земле, но эти представления были скорее гениальными догадками. Лишь в XVIII веке было сформулировано основанное на биологических фактах учение об изменяемости видов животных и растений – трансформизм, – которое противопоставлялось креационизму, утверждающему неизменность видов.
Учение, которое стремится вскрыть причины и закономерности биологической эволюции, возникло благодаря трудам Ж. Б. Ламарка («Философская зоология», 1809) и Ч. Дарвина («Происхождение видов путём естественного отбора», 1859).
Учение Ч. Дарвина об эволюции органического мира основано на триаде: изменчивость, наследственность и отбор. Изменчивость может быть наследственной («неопределённой») и не наследственной («определённой»). Отбор в природе происходит непрерывно. Ч. Дарвин назвал его «естественный отбор». Человек в своей селекционной деятельности применяет кроме того «искусственный отбор». Этой форме отбора он посвятил книгу «Изменение домашних животных и культурных растений». Преследуя разные цели, люди проводили искусственный отбор в различных направлениях. В результате породы дивергировали, т. е. расходились и образовывались новые породы с присущими им признаками. В дикой природе также происходит дивергенция. Отдельные особи определённого вида составляют естественную группировку на отдельном участке ареала данного вида и называются популяцией. Совокупность географических популяций с устойчивыми признаками, способных скрещиваться и давать плодовитое потомство, называются подвидом. При дивергенции могут возникать новые виды, обладающие свойством не давать потомство с другими видами.
 |
|
|
М. Б. Ламарк |
В настоящее время систематики распределяют организмы по группам, используя следующую классификацию: вид, род, семейство, отряд, класс, тип. Существует бинарная номенклатура, упоминается род, затем вид, например: Canis familiaris – собака домашняя, Canis lupus – волк; современный человек – Homo sapiens.
Механизм эволюции Ж. Б. Ламарк и Ч. Дарвин понимали
У Ч. Дарвина о той же шее жирафа сказано следующее: «Жирафа по своему высокому росту, очень длинной шее, передним ногам, голове и языку прекрасно приспособлена к обрыванию листьев верхних ветвей деревьев… Самые высокие особи, которые были на дюйм или на два выше других, могли часто сохраняться в период засухи, бродя в поисках за кормом по всей стране… Если принять во внимание вероятный образ жизни жираф, то те особи, у которых какая-либо или несколько разных частей тела были длиннее, чем обыкновенно, вообще имели преимущество и выживали. При скрещивании они должны были оставлять потомков или с теми же самыми особенностями строения, или с наклонностью изменяться в том же направлении, тогда как особи, организованные менее благоприятно в этом отношении, должны были оказаться наиболее склонны к гибели… Естественный отбор и охраняет и тем самым выделяет всех более высоких особей, давая им полную возможность скрещиваться, и способствует уничтожению всех более низких особей».
 |
|
|
Ч. Дарвин |
Ч. Дарвин оказал глубокое воздействие на развитие всех биологических наук. В начале XX века возникли новые науки: генетика – наука о наследственности, экология – наука о взаимодействии между организмом и средой, селекция – наука об отборе для получения пород животных и сортов растений. Эволюционный метод пронизал систематику животных и растений, палеонтологию, анатомию – где утвердился раздел сравнительной анатомии, также и сравнительной эмбриологии. В физиологии началось историческое изучение животных и человека, преемственности высшей нервной деятельности. Новые направления развились в геоботанике и геозоологии и во многих других биологических науках.
Дарвинизм – не застывшая наука и свой вклад в теорию эволюции внесли многие учёные. Здесь нет времени упоминать всех. Назовём русских учёных: В. О. и А. О. Ковалевские, К. А. Тимирязев, И. П. Павлов, Н. И. Вавилов, А. Н. Северцев, С. С. Четвериков, И. И. Шмальгаузен. Вся генетика пронизана представлениями о наследственности, изменчивости и отборе. Крупные отечественные генетики сделали многое в области теории эволюции: Н. П. Дубинин – автогенетический процесс, Н. В. Тимофеев – способствовал внедрению математики в генетику, И. А. Рапопорт – пионер химического мутагенеза, Л. Н. Делоне – пионер радиоселекции (вслед за открытием X. Мёллера радиомутабильности у дрозофилы), И. И. Шмальгаузен – стабилизирующий отбор, Д. К. Беляев – дестабилизирующий отбор, Б. Л. Астауров, Н. А. Струнников – практическое применение андрогенеза и полиплоидии. Нужно сказать, что эволюционная теория в начале прошлого века получила большой приток исследователей, пока научная среда не развалилась под натиском Т. Д. Лысенко, которого поддерживал Сталин. До сих пор биологическая наука не полностью оправилась от нанесённого ущерба. Тем более отвратительна та путаница понятий, которая вносится недоучками в эволюционную теорию в наше время.
Создание всякой научной теории формируется в борьбе суждений, однако дискуссии вокруг целостной теории эволюции принимают часто уродливые формы, и высказываемые оппонентами этой теории суждения нередко выдают полное незнание биологии.
 |
|
|
И. И. Шмальгаузен |
Кроме ламаркизма, существует ряд концепций, отрицающих значение отбора: автогенез, мутационизм, номогенез и пр. В данной статье мы не будем разбирать эти представления за недостаточностью места. Кроме того, хотя эволюция свойственна всей живой природе, мы не будем говорить об особенностях эволюции прокариот, в клетках которых нет ядра, а только эукариот, имеющих оформленное ядро в клетке.
Эволюционные процессы, протекающие внутри вида и завершающиеся видообразованием, называют микроэволюцией, развитие группы таксонов надвидового ранга – макроэволюцией. Отбор происходит во времени и в пространстве и идёт по фенотипу (сумма признаков) а не по генотипу (сумма генов). Именно поэтому отбор играет тактическую, а не стратегическую роль, и, следовательно, не может идти по логике «движущей силы к совершенству». В природе существует принцип подстраховки, а не экономии. Благодаря этому всегда есть в геноме запас «молчащих», «немых» генов, которые могут быть использованы при выходе из возможного тупика. Природе свойственна многомерность ходов развития.
В наше время появилось много исследований с поисками близости и удалённости различных таксонов по различным критериям. Наибольшее число таких работ посвящено анализу ДНК у различных объектов. Вышла великолепно изданная книга «Программа № 25 фундаментальных исследований Президиума Российской академии наук "Происхождение и эволюция биосферы"» (Новосибирск, 2006). Исследований с привлечением морфологических, анатомических, физиологических и др. методов проводилось много и они безусловно полезны. Ещё в 1915 году Л. Н. Делоне предложил метод кариосистематики. Проведя сравнительно-кариологические исследования видов Muscari, он отнёс два вида в другой род, а именно Bellevalia. При всей необходимости работ по сравнительной таксономии с выбором различных критериев для сравнения вплоть до молекулярных, наше время требует углублённого и честного проникновения в синтетическую теорию эволюции. Тем более, что всё больше появляется обескураживающих ревизий дарвинизма.
Уже сейчас есть заделы для дальнейшего развития теории эволюции. Нельзя «закапываться» в сиюминутные задачи и тратить время на унизительные дискуссии с модными невеждами. Следует работать «на опережение». Нужно формулировать проблемы будущего, не только основываясь на интуиции, но и опираясь на тот научный багаж, который уже накоплен, но никак не используется.
За последнее время наиболее значительным новым представлением является «синергетика» И. Р. Пригожина. В книге Пригожина и И. Стенгерс «Порядок из хаоса. Новый диалог человека с природой» (КомКнига, 2005) дана теория диссипатичных структур для неравновесных открытых систем, которая в общей форме приложима к принципиальным закономерностям, играющим роль в биологической эволюции.
I. Проблема резерва наследственности. Молчащие гены. Немые гены
 |
|
|
Н. И. Вавилов |
Для каждого вида высших организмов присуще большое число генов, которые считываются в течение онтогенеза, причём в каждой дифференцированной клетке считывается только незначительная часть генов генома, остальные заблокированы и носят название «молчащих». Именно смена «работы» одних кластеров генов другими даёт возможность в многоклеточном организме иметь разную специфическую функцию каждой клетки, т. е. дифференцироваться, несмотря на то, что геномы всех этих клеток в большей степени одинаковы.
Однако, экстремальные факторы могут вызвать взрыв изменчивости. Происходит дестабилизация кариона (клеточного ядра), когда начинают считываться гены, которые не характеризуют совокупность генов данного вида, но передаются по наследству. Это резервный пул генов. Мы назвали эти гены «немыми» (Н. Л. Делоне. Человек, Земля, Вселенная. М., 2004). Резерв «немых» генов даёт возможность новообразованию видов. Только благодаря наличию немых генов состоялась возможность появления второплавающих животных, атавизмов, одомашнивание животных. Виды длительное время остаются стабильными, что определяется постоянством их геномов, но существует пул немых генов, откуда как из кладовой при ускорении эволюции в периоды катаклизмов «черпаются» новые работающие гены.
Следовательно, генетические возможности живых организмов Земли способны обеспечить эволюцию видов в космических поселениях за счёт взрыва изменчивости, а также ускорение эволюции при катаклизмах на Земле.
II. Проблема изменчивости
Изменчивость эукариот (организмом, имеющим клеточное ядро) может происходить благодаря различным причинам:
1. Изменение структуры гена – в результате мутаций внутри гена.
2. Изменение дозы гена – как следствие уменьшения или увеличения числа одного и того же гена. Причиной служат мутации типа делеций и дупликаций при хромосомных перестройках, при политении и уменьшение и увеличение числа хромосом в наборе при полиплоидии ианэуплоидии.
3. Дестабилизация кариона – при этом происходят изменения в отношениях эухроматин-гетерохроматических блоков в хромосомах, что может быть вызвано внешними условиями, а также при перестройках хромосом типа инверсий и транслокаций, приводящих к изменениям места кластеров генов и соседних с ними вставках гетерохроматина по длине хромосомы. Эухроматические участки содержат основное число генов. Гетерохроматические участки играют роль в функционировании генов. Дестабилизация кариона приводит к перемене положения гена в архитектонике ядра, что отражается на регуляции целых кластеров генов, перевод молчащих генов в состояние считывания информации и наоборот.
4. Перекомбинация генов – благодаря гибридизации. При этом целый гаплоидный набор отцовских хромосом соединяется с материнским, образуя новый диплоидный набор зиготы.
5. Перенос отдельных генов – при генной инженерии, причём возможен перенос генов очень отдалённых таксонов.
III. Проблема фенотипической адаптационной изменчивости
Ненаследственная изменчивость, модификации, может быть разной природы. Она может происходить в результате мутаций в соматических клетках, тогда такие нарушения не будут наследоваться, поскольку гаметы останутся в норме. Также модификации могут приводить к уродствам. Но большинство ненаследственных изменений носят характер фенотипической адаптации, т. е. приспособления к условиям среды. В данном случае большую роль играет «доза гена».
Один и тот же состав генов может
Признак формируется сложно. От гена до признака длинный путь. Признаков, появляющихся в результате деятельности одного гена, значительно меньше, чем признаков, зависящих от совокупности ряда генов, которые считываются в каждый данный определённый момент. Формирование признаков в онтогенезе – это результат цепи процессов и их регуляции.
Гены в геноме – это не кнопки на приборной доске: сколько кнопок нажмёшь – столько лампочек включится.
Роль генов в фенотипе идёт не в виде команды
Совокупность генов – чрезвычайно существенное звено в общей цепи событий, происходящих в клетке. Однако, геном испытывает обратное влияние со стороны организма, и такая обратная связь знаменует создание организма как целого.
|
Искусственный отбор привёл к разнообразию пород собак: |
||||||
 |
 |
 |
 |
|||
Волк |
ирландский терьер Рона |
сенбернар Пьер |
болонка Мася |
|||
IV. Проблема отбора
Отбор в популяциях идёт не по генному составу, а по признакам организма. Фенотип играет основную роль. В геноме могут резервироваться возможности, которые в определённое время проявляются в фенотипе. Есть много форм отбора: естественный, искусственный, массовый, индивидуальный, внутрилинейный, центростремительный, центробежный, деструктивный, однолинейный, нормализующий, периодический и др. Именно понятия о естественном отборе, происходящем в дикой природе, и искусственном отборе, проводимом человеком в его хозяйственной деятельности – являются краеугольным камнем дарвинизма.
 |
|
|
Д. К. Беляев |
Рассмотрим две формы отбора: стабилизирующий и дестабилизирующий. «Стабилизирующий отбор» – понятие, которое ввел И. И. Шмальгаузен (1968). При этом отборе происходит элиминация факторов, непосредственно зависящих от наличия гена (генов) с неблагоприятным действием и отличающихся от популяционного среднего, типичного для данного генофонда.
Представление о «дестабилизирующем отборе» принадлежит Д. И. Беляеву. Он расширил представление об отборе, вводя одновременно в это понятие как увеличение изменчивости за счёт числа активных генов в геноме, так и особенности отбора: «Дестабилизирующий отбор в кратчайшие сроки ломает систему онтогенетической регуляции признаков и функций, сложившихся под действием стабилизирующего отбора, и порождает громадный размах изменчивости» (1981).
Например, в особых условиях космического полёта в невесомости создадутся условия для дестабилизирующего отбора. Даже в тех полётах в Космос, которые проходят сейчас (со сравнительно небольшой длительностью): «Необходимо подчеркнуть, что формирующиеся в процессе адаптации к невесомости сдвиги в организме человека и животных адекватны новым условиям существования» (А. И. Григорьев, А. Д. Егоров, 1998). В длительных полётах будет ломаться система онтогенетической регуляции признаков и функций, сложившихся под действием стабилизирующего отбора, и следует ожидать увеличения изменчивости. Вместе с тем отбор будет идти в этих же новых условиях и, безусловно, примет особое направление, отличающееся от стабилизирующего в земных условиях.
Изучен механизм передачи информации от структурного гена до первичного белка. Это, однако, не означает, что поток информации от гена к признаку прослеживается в организмах живых существ. Сложный комплекс взаимодействий приводит к дифференцировке клеток, а самосборка отдельных конструкций во время онтогенеза играет особую роль в становлении целостного организма. В пространстве и во времени возникают сложные сети взаимоотношений, они подвижны и уровни регуляции идущих в целостном организме процессов многообразны и многоканальны.
Теории для понимания общего нельзя строить на экзотических исключениях. Всем известен пример, когда в гене, который кодирует фенилаланингидроксилазу, возникает мутация, приводящая к отсутствию в организме этого фермента – и возникает болезнь фенилкетонурия. Можно и дальше приводить подобные примеры, но это только исключения. Именно исключением можно назвать случаи, когда отдельный ген, кодирующий один первичный белок, приводит к появлению признака. В медицинской генетике менделирующих болезней изучено меньше, чем так называемых полифакториальных наследственных заболеваний. Хотя по Менделю каждый ген представлен двумя аллелями, приводящим к различным признакам, и гетерозиготное носительство может содержать опасность. Что касается полифакториальных наследственных заболеваний, то это просто «белое пятно» на карте.
В последнее время появилось много высказываний об отсутствии роли отбора в эволюции. Мода даже
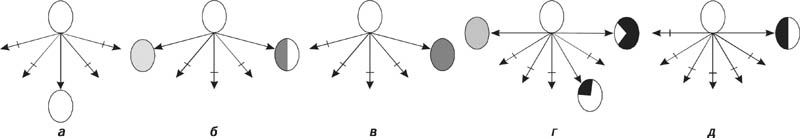 |
|
Формы отбора в популяциях: |
V. Проблема формирования популяций в поселениях вне Земли
Космическая эра началась, и придёт её новый этап, когда будут образованы космические поселения, и поколения за поколениями люди будут жить в невесомости. Это время безусловно наступит.
Процесс формирования популяций и их динамика станут предметом изучения. Популяция формируется за счёт изменчивости и
отбора. В невесомости они могут происходить иным образом, чем на Земле. Адаптация частично обусловлена влиянием
среды, но определяется главным образом генетически. Полимерные гены, которые в основном участвуют в создании
количественных признаков, создают широкие возможности для полиморфизма. Выжить и размножаться могут только те особи,
генотип которых даёт им
Чрезвычайную роль играет величина популяции. В малых популяциях наступает действие генетико-автоматических процессов со случайным дрейфом генов, что нужно иметь в виду, рассчитывая численность особей в популяции. Генетико-популяционный аспект будет иметь большое значение при формировании космических поселений.
Рассмотрим космические популяции человека с позиций менделевской генетики. По классическому определению:
популяция – это группа свободно скрещивающихся или потенциально способных к скрещиванию особей, населяющих
определённый ареал и обнаруживающих определённые пространственно-временные взаимоотношения. Теоретический анализ
менделевской популяции, проведённый независимо друг от друга Харди и Вайнбергом, показал, что в любой панмиксной
популяции имеется закономерное распределение генотипов. Если в популяции наследуются аллели любого гена, например,
доминантная аллель «А» и рецессивная аллель «а», они представлены определёнными концентрациями, под которыми
понимается относительная доля данного аллеля от их суммы в популяции. Расчёт производят по формуле
 |
|
|
Г. И. Мендель |
Естественный отбор как категория, ведущая эволюцию, потеряла своё первенствующее значение для человека. Социальные
закономерности определяют структуру человеческого общества. Однако, это не значит, что у человека не действует отбор
отрицательных мутаций, вызывающих уродства и гибель в раннем возрасте, гибель в половозрелости, раннюю смертность
и др. Человеческие популяции отягощены врождёнными наследственными заболеваниями: до 4 % детей в каждом
поколении рождаются поражёнными отрицательной наследственностью, полученной от их родителей, которая у них была скрыта
Отбор вмешивается в распределение генов по разным популяциям людей, живущих в разных условиях среды. Для понимания
основ биологической природы человека генетика популяций и анализ деятельности отбора имеют огромное значение. Наиболее
простой пример – это изменение концентрации аллелей в популяции, изменяющих соотношения, которые соответствуют
формуле Харди – Вайнберга. Если отбор направлен против рецессивных гомозигот «аа» (например, если «аа» составляют
99 потомков по сравнению со 100 потомками от особей с генотипами «АА» и «Аа»), то в каждом последующем
поколении концентрация аллеля «А» будет увеличиваться, а концентрация аллеля «а» – уменьшаться.
Отрицательная селекция (S) против генотипа «аа» составляет величину 0,01, в то время как для
генотипов «АА» и «Аа» –
В популяциях могут наблюдаться колебания числа особей, образуя так называемые «популяционные волны».
Величина популяций может уменьшаться до минимума, и только восстановление величины популяции уменьшает опасность её случайного вымирания. Критическая величина популяций различна для разных степеней интенсивного отбора, поэтому определённая величина популяций для одних генов может ещё не достигнуть критической, в то время как для других этот предел уже превышен.
Число особей в популяции зависит от территории, которую занимает популяция: может наступить эффект «популяционного давления», когда фактическое число особей в биотике превышает число, которое может вместить соответствующее жизненное пространство.
 |
|
|
К. Э. Циолковский |
В ограниченных или малых популяциях осуществляется случайная дифференцировка по разным аллелям. Эти стохастические
процессы изменений в концентрациях аллелей называют «генетико-автоматическими процессами» или «дрейфом генов».
Появились специфические сочетания разных аллелей в поселениях, получивших начало от первых колонистов. Есть область
генетики, которая занимается «изолятами», т. е. отдельными, далеко расположенными поселениями. Иногда в таких
поселениях хорошо проявляется «эффект родоначальника». Это бывает, если у
Итак, какое наименьшее число пар может осуществить тот коллектив, который послужит основой внеземной колонии?
Представляется, что в данном случае можно взять в качестве модели Ноев ковчег. Ной имел трёх сыновей и трёх невесток.
Три пары могут оказаться достаточными для поставленной задачи. Конечно, при этом эти три пары должны рожать здоровых
детей. Да и в дальнейшем может их ожидать множество случайностей. Например, самое простое, можно потерять
Менделевские закономерности не объясняют распределения по территории многих количественных признаков. Примеров этому можно было бы привести большое число. Очень яркий пример – это виды , имеющие добавочные хромосомы. Особей, обладающих добавочными хромосомами, всегда оказывается больше по краям ареала данного вида. Совершенно новые условия космических поселений будут вызывать дестабилизирующие процессы, которые наложат свой отпечаток на закон Харди – Вайнберга. Превалирующую роль займёт дестабилизация кариона. Эволюция будет проходить с ускорением. Произойдёт evolution explosive – эволюционный взрыв.
VI. Проблема целостности биосферы
Биосфера состоит из литосферы, гидросферы и атмосферы, пронизанных живыми существами, которые определяют круговорот веществ. Создал представление о биосфере В. И. Вернадский в 1923 г., хотя термин был предложен ещё в 1875 г. Э. Зюссом.
 |
|
|
В. И. Вернадский |
Биосфера Земли имеет длительный срок развития. Эта сфера жизни подвергалась катаклизмам, разрушалась и воссоздавалась во всё более совершенной форме. Не только животный мир менялся, но и химический состав биосферы претерпевал изменения.
Воздействие человеческой цивилизации на биосферу В. И. Вернадский характеризовал так: «Человек в этой форме
своей жизни является геологической силой большего значения, чем мы это себе представляем. И эта сила, созданная
всей предшествующей жизнью, не может ни исчезнуть, ни повернуть назад»… «Изменение лика природы, созданное культурной
средой жизни, есть такой же стихийный процесс, как любое проявление геологических сил нашей планеты – и столь же
мало может быть изменено». Та тревога, которую нельзя не ощущать, видя последствия антропогенной деятельности на
биосферу, приводит к прекраснодушным призывам вернуться в патриархальное прошлое. Но цивилизация
Человечество стало наносить всё более и более глубокие «пробоины» в круговорот природы. И может быть самые чувствительные раны наносятся духовному миру, а души начинают черстветь. Пора спохватиться, перестать, захлебываясь, мчаться вперёд, развивая технику и начать чинить «прорехи» в биосфере и своей душе. Ничего нет более кощунственного, чем лозунг: «Победить природу – наша задача!». Однако, значительно чаще без всякой идеи, просто толкаемые алчностью, «сиюминутные люди» подрывают возможность продолжения жизни на Земле.
Возможно, что мечта о космических поселениях людей – это подсознательное желание «начать жить заново». Жить лучше, правильнее и не делать ошибок. Сразу же возникает вопрос: возможно ли воссоздать биосферу с круговоротом веществ? Ведь это замкнутая самообновляющаяся система. Конечно, размеры системы очень велики – это «плёнка Земли», но она самосохраняется в своём развитии. Даже теплицы для растений требуют много «протезов». Искусственное очищение от отходов в первую очередь. Но ведь необходимо, чтобы микроорганизмы и растения в теплице сохраняли возможность взаимополезного сосуществования. Самое главное – атмосфера в такой теплице, её состав; очищение почвы и её регенерация. Задачи, которые стоят перед построением таких теплиц, требуют очень большого внеприродного, механического обслуживания и разнообразных биологических добавок. Такое устройство на «протезах» – это не прообраз биосферы.
Пока люди прикреплены «пуповиной» к своей Земле. Но не хочется быть скептиком. Во всяком случае, построение космических поселений с авральным посещением с Земли и возвращением опять на Землю вполне реально. Мечты о Космосе должны пробудить в людях желание беречь и сохранять биосферу нашей Земли.
Заключение
Трудно представить, что сейчас делается необходимым защищать общебиологическую теорию, поскольку на эволюционное учение идёт вал бесцеремонной, неприкрытой глупости. Убогий лепет, полное незнание теории эволюции выдаётся за праведную борьбу с дарвинизмом. Но ведь это не ново, была уже «обезьянья война» в
* Разумеется, опечатка первоисточника – имеется в виду 1859 г. XIX века. (Примеч. вед. сайт «Разум или вера?» от 10.05.2026 г.)
|
|
|
|